
Tous droits réservés pour tous pays. All rights reserved.
| Serveur © IRCAM - CENTRE POMPIDOU 1996-2005. Tous droits réservés pour tous pays. All rights reserved. |
Journal of the Acoustical Society of America, 22 March 1996
Copyright © ASA 1996
a)Some of the experiments reported here were conducted in partial fulfillment of the requirements for C.M.H. Marin's (1991) doctoral dissertation which is available in French.
Polyperiodicity (Marin, 1991),
or the presence of several periodic sounds with different periods that are not
integer multiples of one another, has also been tested by several
investigators. Polyperiodicity is usually produced experimentally by
introducing a difference between the fundamental frequencies (F0) of two or
more harmonic complexes. The identification of two simultaneous vowels improves
as the difference between their respective F0s ( F0) increases from
zero to one or two semitones (6-12%) (e.g. Scheffers, 1983; see de Cheveigné et al., 1995 for a review). However, Chalikia and Bregman
(1989)
found that segregation was more difficult for a F0 of an octave than
for a separation of 0.5 or 6 semitones. This result suggests that it is not
simply F0, but perhaps polyperiodicity or harmonic non-coincidence (de Cheveigné, 1993)
that is responsible for the phenomenon: the combination of two harmonic sounds
separated by an octave gives a waveform with a single period. Indeed, work by
Marin (1995)
is consistent with this hypothesis.
F0) increases from
zero to one or two semitones (6-12%) (e.g. Scheffers, 1983; see de Cheveigné et al., 1995 for a review). However, Chalikia and Bregman
(1989)
found that segregation was more difficult for a F0 of an octave than
for a separation of 0.5 or 6 semitones. This result suggests that it is not
simply F0, but perhaps polyperiodicity or harmonic non-coincidence (de Cheveigné, 1993)
that is responsible for the phenomenon: the combination of two harmonic sounds
separated by an octave gives a waveform with a single period. Indeed, work by
Marin (1995)
is consistent with this hypothesis.
The research reported here studied the possibility that F0 and FM may
give rise to other auditory cues that can be used in situations in which
listeners are trying to decide whether or not a target sound with known
properties is embedded in a complex background sound. Two of the experiments
reported below (Exps. 1 and 4) formed part of a larger series of experiments
that employed a task in which subjects were to match one of two test formants
embedded within background sounds to an isolated target formant
(Marin, 1991).
These two experiments were concerned with the segregation cues frequency
modulation and polyperiodicity, respectively. FM may reduce to polyperiodicity
in stimuli with few components in which those with different FM patterns do not
stimulate the same auditory filters. Further, polyperiodicity is a potent cue
for perceptual segregation of concurrent harmonic sounds. However, both cues
may also provide important temporal information that can be used by top-down
selection processes in deciding whether or not a given target sound is likely
to be embedded in a given background sound even in cases where that sound is
not fully segregated. Indeed, while conducting Experiments 1 and 4 of the
present series, it was noted that for both cues, the components of the test
formant and background sounds interacted with one another and created auditory
beats, or roughness. The research presented here investigated the contribution
to target matching performance of amplitude envelope fluctuations produced by
these parameters. Accordingly, four additional experiments were
performed. Experiments 2 and 3 studied frequency modulation in stimuli in which
beats were considerably reduced. Experiment 5 investigated the polyperiodicity
cue with stimulus conditions similar to those in Experiment 3. In addition,
computer simulations of peripheral auditory filtering were performed to model
the degree to which auditory beating was present in the different stimulus
conditions investigated. Finally, Experiment 6 sought to determine the
amplitude modulation depth at which subjects were able to detect the frequency
region in which the interactions occurred when AM was the only available cue.
These latter results are then used to evaluate the potential contribution of
envelope fluctuations in the five other experiments.
F0 cues, the backgrounds were designed
to have spectral envelopes that were the inverse of those for their respective
test formants. Both test formants and background sounds were derived from a
harmonic series with a nominal F0 of 250 Hz.
The formants used in Experiments 2, 3, 5, and 6 were "flat" formants composed of two or three equal-amplitude harmonics. Their center frequencies were 375, 750, 1250, and 1750 Hz. These center frequencies were chosen to be close to those of the "normal" formants. The 375 Hz formant was the only one to have two components and its CF was the mean of the frequencies of the first two harmonics.
The backgrounds for the "flat" formants were designed in the same way. They were composed of the harmonic series with components situated on each side of the components of the flat formant, but were completely missing the components corresponding to those of the formant.
With the exception of stimuli for Experiment 6, sound files were synthesized with the Csound program (Vercoe, 1986) on a VAX 11/780. An additive synthesis algorithm was used in which each frequency component was generated by a digital oscillator whose amplitude and frequency were controlled in a continuous fashion. The spectral envelopes for the stimuli of Experiments 1 and 4 were stored as interpolated table look-up functions, the amplitude of each component being determined by its instantaneous frequency. The stimulus waveforms were digitally synthesized at a sampling rate of 10 kHz. All calculations took place in 32-bit floating-point format and the waveform was then stored in 16-bit integer format on disk. Each stimulus was transferred to the hard disk of a Macintosh II which controlled the experiment.
For Experiment 6, stimuli were synthesized in real-time at a sampling rate of 12 kHz with a DSP card (Smith and Chervin, 1986) controlled by the Macintosh II. Calculations took place in 24-bit integer format and were output in 16-bit integer format.
In all experiments, digital waveforms were converted to analog signals through Burr-Brown 706 DACs. The signal was passed through two Rockland 432 low-pass filters in series, each giving -48 dB/oct attenuation. The cut-off frequencies were set at 40% of the sampling rate. The filtered signal was amplified with an MB Systems 105a power amplifier and presented diotically over Beyer DT48 earphones at a level of 75 dBA (Experiments 1, 2, 4, and 6) or 59 dBA (Experiments 3 and 5). To verify the presentation level, each earphone was connected via a flat-plate coupler to a Bruel and Kjaer 1209 sound level meter (A-weighting). The subject was seated in a Soluna SN1 double-walled sound attenuation booth during the experiment.
During early testing, it became apparent that different subjects did not have the same thresholds. In order to avoid experimental runs that were too long, due to a small step size or a large range for the adaptive procedure, these parameters were fixed separately for each subject in each experiment and are reported below. The tracking procedure started at the specified maximum value and remained there until the subject made a correct response. When a subject's tracking stayed too close to the top or bottom of the range, the run was rejected and the values were readjusted.
Within each experimental run, the trials composing the tracking procedure for the different target formants were interleaved in a random fashion. If threshold was determined for a given target before that of the others, trials containing that target at the threshold value continued to be presented, though further data were not recorded for it.
At the beginning of each run, subjects were allowed to listen at will to pairs of stimuli that were to be presented in the experiment, so that they could easily associate the isolated target with the test formant of identical CF embedded in the background and distinguish it from the embedded test formant with a higher CF. The target CF could be changed by pressing a button. These pairs were presented at the highest value of the dependent variable that had been fixed for that subject.
A strong learning effect was observed at the outset of testing. Therefore subjects were trained for several runs until their thresholds appeared to stabilize. Only the last two (Exps. 1-3) or five (Exps. 4-6) runs for each experiment were retained for further analysis.
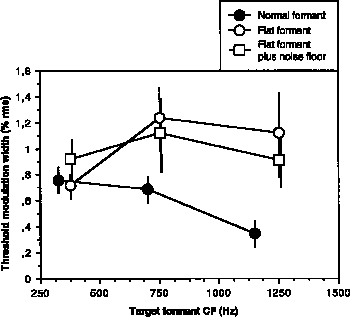
Figure 1. Summary of frequency modulation results for Experiments 1-3 (normal formants, flat formants, and flat formants plus noise floor, respectively). Mean target matching thresholds, expressed as percent rms modulation width, are plotted as a function of target formant CF. Some means do not include values for subjects unable to attain threshold (see text). Vertical bars represent +/- one standard error of the mean.
The thresholds in this experiment were higher than the mean frequency modulation detection thresholds (FMT) found in studies that did not have embedded components: 0.09% rms by Hartmann and Klein (1981) and 0.18% rms by McAdams (1984, App. D) Some thresholds, however, were lower than the mean 0.44% rms threshold measured by Demany and Semal (1989). Thus, with one possible exception, the FM should have been easily detectable at matching threshold, though detectability is clearly not sufficient to perform the present task.
The matching thresholds found here are roughly equivalent to detection thresholds found for a frequency modulation incoherence discrimination task by McAdams and Marin (1990) for similar multi-component targets. The task in that experiment was to detect which of two intervals contained incoherent, aperiodic modulation functions on two subsets of components. In the earlier study however, thresholds increased for higher CF targets rather than decreasing as in the present experiment. The thresholds of the current experiment are smaller by a factor of 3 than Carlyon's (1991, Exp. 4) thresholds for discrimination of sinusoidal FM incoherence due to a modulation phase delay on the middle component of a three-component complex.
It is likely that the perception of low-rate beating induced by frequency modulation in this experiment was a contributing factor in target formant matching by allowing subjects to detect the frequency region where the formant was situated. Low-rate beating has several sources in these stimuli, three of which include:
1) Within-channel interactions between nearby test and background components, the beat frequency of which would vary over time as the components converged, crossed and diverged again. For a given difference between the fundamental frequencies of test formant and background, the beat rate would be higher for higher CF formants since the maximal frequency difference is proportional to the harmonic rank.
2) Amplitude modulation induced by tracing the formant spectral envelope, which would have the greatest modulation depth on the flanks of the formant peak.
3) Amplitude modulation induced by components situated in the skirts of auditory filters.
The first two of these sources can be reduced by replacing the second-order "normal" formants by two- or three-component "flat" formants. The frequency separation between test and background components is thus increased and component amplitudes no longer change with F0 (i.e. envelope tracing is eliminated). This modification was studied in Experiment 2.
In order to compare the data from Experiments 1 and 2 (Fig. 1, filled vs. open circles), an analysis of variance with factors formant type (normal vs flat formants), target formant CF, and repetitions were performed on the mean matching thresholds for all subjects (with missing data for two subjects in Experiment 2). The main effect of formant type was highly significant [F(1,80) = 9.95, p<0.005] and the interaction between formant type and target formant CF just attained significance [F(2,80)=3.14, p<0.05]. Planned comparisons between experiments for each formant CF showed no difference between the 325 Hz formant and the 350 Hz flat formant [F(1,80)=0.029, n.s.], a significant difference between the 700 Hz formant and the 750 Hz flat formant [F(1,80)=5.77, p<0.05], and a highly significant difference between the 1150 Hz formant and the 1250 Hz flat formant [F(1,80)=10.58, p<0.005].
Interactions between adjacent components of the modulated test formant and unmodulated background that stimulated the same auditory channels may have persisted in the stimuli of the present experiment, particularly for the higher CF formants. They would have a relatively high rate around 250 Hz with a superimposed 5 Hz periodicity due to the varying mistuning of the harmonic ratios by the FM. In the auditory channels in the region of the modulated test components, a further cue could be fluctuations in activity induced by the back and forth motion of the excitation envelope. The lack of change of thresholds for the lowest CF target suggests that envelope tracing and local component beating did not contribute in the first place at this CF or that another, equally potent, cue, such as AM induced by the auditory filters, was used. The contribution of this source of AM can be evaluated by comparing the FM data with those obtained using a static mistuning (see section III).
In the stimuli of the present experiment, beats may also have been caused by the presence of combination tones (CT) (Goldstein, 1967; Plomp, 1976): nonlinear distortion products created by the background components may have interacted with the test formant components and vice versa. One can eliminate the difference tone (f2-f1) by presenting the stimuli at a spectrum level below 50 dB SPL. The level of the most intense higher-order CT (2f1-f2), when f1 and f2 are adjacent components, is less than that of either component by about 15 dB and could thus be masked by a noise floor (Plomp, 1976). Further, given the level difference between stimulus components and possible CTs, any beating produced would be very weak. The possible further reduction of beats between partially resolved test and background components and between stimulus components and CTs by the introduction of a noise floor was investigated in Experiment 3.
A comparison of the data for Experiments 2 and 3 (Fig. 1, open circles and squares) revealed that the introduction of a noise floor did not increase thresholds. An analysis of variance (with missing data for two subjects from Experiment 2 and one from Experiment 3) on factors stimulus type (with or without noise floor), target formant (3 CFs), and repetitions (2) confirmed that neither the main effect of stimulus type nor its interaction with formant CF were significant [F(1,78) = 0.07, F(2,78)=0.52, respectively].
The possibility that cues such as AM induced by auditory filter skirts resulting from frequency modulating the test formant components contributed to target matching can be partially evaluated by comparing these data with those obtained from similar stimuli in which a static mistuning is applied to the components of the test formant. This stimulus modification was performed in Experiments 4 and 5.
Certain modifications were made to shorten the procedure in this experiment. Only 12 turnarounds were recorded in the adaptive procedure, the last eight being used to compute the threshold. The step size in the first two turnarounds was four times the final one and that in the second two turnarounds was two times the final one. The final stepsize remained constant for the last eight turnarounds. In addition, only the isolated target CFs of 325 and 1150 Hz were studied. Subjects began by doing six runs. The results of the first run were not analyzed. If the interval between the highest and lowest thresholds over five consecutive runs was greater than a criterion value of 1.5 times the final stepsize, the subject was required to complete additional runs until this criterion was met. The analysis was performed on the last five runs.
The starting (maximum) values of F0 used in the adaptive procedure
were 0.2% and 2.0%. The stepsizes were 0.01% and 0.1%. For four subjects, the
lower values were used for the 1150 Hz formant and the higher values for the
325 Hz formant. For one subject the lower value was used for both formants.
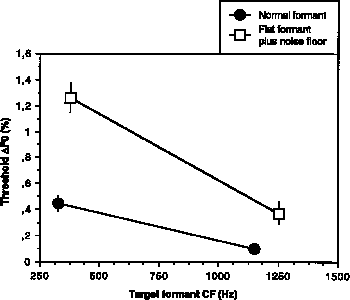
Figure 2. Summary of polyperiodicity results for Experiments 4-5 (normal formants and flat formants plus noise floor, respectively). Mean target matching thresholds, expressed as percent difference in fundamental frequency, are plotted as a function of target formant CF. Vertical bars represent +/- one standard error of the mean.
F0 found by other studies to be necessary to
give improved identification of embedded vowels
(Chalikia and Bregman, 1989; Culling and Darwin, 1993; Cutting, 1976; de Cheveigné
et al., 1995; Gardner and Darwin, 1986; Lea, 1992; Scheffers, 1983; Summerfield
and Assmann, 1991).
This sizable difference in threshold may be related to the task demands. In the
studies cited, subjects were required to identify vowels. In our experiment,
with presentation of an isolated target formant immediately prior to the test
stimulus, the decision concerning which test stimulus contained a formant
matching the target would have to have been based on (most likely timbral) cues
that signaled the frequency region of the test formant. These cues were quite
likely derived from beats created in the region of the test components by
interactions with background components. It should be noted that in the absence
of the background, this task is trivial. However, even unusually small
mistunings can apparently create sufficient fined-grained temporal interactions
to perform the task, especially when the component spacing is small on the
basilar membrane as is the case with the higher CF formant. This effect may
result from interactions similar to those described for mistuned harmonics by
Moore et al.
(1985) and Hartmann et al.
(1990).
It is interesting to compare the thresholds for this experiment with those
obtained in Experiment 1. Assimilating rms modulation thresholds measured in
Experiment 1 and the F0 thresholds from Experiment 2 to the sam
dependent variable, a repeated measures ANOVA was performed on the mean data
across repetitions for the 5 subjects that completed both experiments, with
factors cue type (FM vs F0) and target formant (325 vs 1150 Hz). The
effect of target CF was significant [F(1,4)=32.2, p<0.005], but neither cue
type nor its interaction with target CF were significant [F(1,4)=2.4 and
F(1,4)=0.7, respectively].1
For the F0 stimuli of Experiment 4, the beat rate between adjacent
components of test formant and background would be equal to the local frequency
difference between the interacting components. The amplitude modulation depth
created by their interaction would be a function of their relative amplitudes,
being greatest if their amplitudes were equal and decreasing monotonically as a
function of their difference. In order to evaluate the potential contribution
of these interactions to target matching performance, the frequency difference
between adjacent components of test and background components was increased and
pink noise was added in Experiment 5.
F0 used in the adaptive procedure
ranged from 0.2% to 8%. The stepsizes ranged from 0.01% to 0.4%. For some
subjects the maximum values and stepsizes were different for the two target
formants.
An unpaired t-test on five threshold estimates for each subject (removing outlying data for one subject at the higher CF target2) showed that the difference between mean thresholds for the two formants was highly significant [t(43)=6.12, p<0.0001].
All subjects' thresholds obtained in this experiment were higher than those
from Experiment 4 using normal formants and no masking noise. An analysis of
variance on factors stimulus type (2), target formant (2) and repetitions (5)
(with outlying data removed for one subject as before) confirmed that the
F0 thresholds were lower for normal formants than for flat formants in
noise [F(1,75) = 46.02; p<0.0001]. However, the interaction between stimulus
type and target formant CF was also significant [F(1,75)=11.28, p<0.005],
indicating that the decrease in threshold that accompanied an increase in
target CF was greater for the flat formant stimuli with the noise floor than
for the normal formants in quiet. Further, the greater reduction of beating
between nearby test and background components for the lower CF target was
probably responsible for the larger threshold increase at that CF (0.8%
increase) than at the higher CF (0.3% increase).
To compare the results for FM and F0 stimuli, the mean thresholds
across repetitions for the two target formants and five subjects common to
Experiments 1, 3, 4, and 5 were submitted to a three-way analysis of variance
(with missing data in Exp. 3 and outlying data removed in Exp. 5 as before).
Thresholds for FM stimuli were expressed as rms modulation width. The
independent variables were cue type (FM vs F0), stimulus type (normal formant in quiet vs flat formant in pink noise), and target formant CF (325/375 vs 1150/1250). The main effect of cue type was not significant [F(1,31) = 0.18]. The main effect of stimulus type was significant [F(1,31) = 11.31,
p<0.005], reflecting the fact that the thresholds for normal formants in
quiet are lower than those for flat formants presented in noise across both cue
types. From comparisons of differences between thresholds for the two stimulus
types, it appears that the use of flat formants against a pink-noise floor
impairs target matching performance more for statically mistuned test formants
than for test formants with frequency modulation at the low CF target
[t(4)=3.6, p<0.05]. At the higher CF target there is no difference between
FM and F0 stimuli [t(3)=0.7, n.s., missing data for one subject].
The larger effect of changing from normal formants in quiet to flat formants in
pink noise for F0 stimuli compared to FM stimuli at the lower CF
suggests a difference in the form of beating created by the two classes of
cues. To examine the nature of beating present in the stimuli for the first
five experiments, and to estimate the relative amount present under the
different experimental conditions, we performed a computer simulation of
auditory filtering and devised a measure of within-channel fluctuation of
activity.
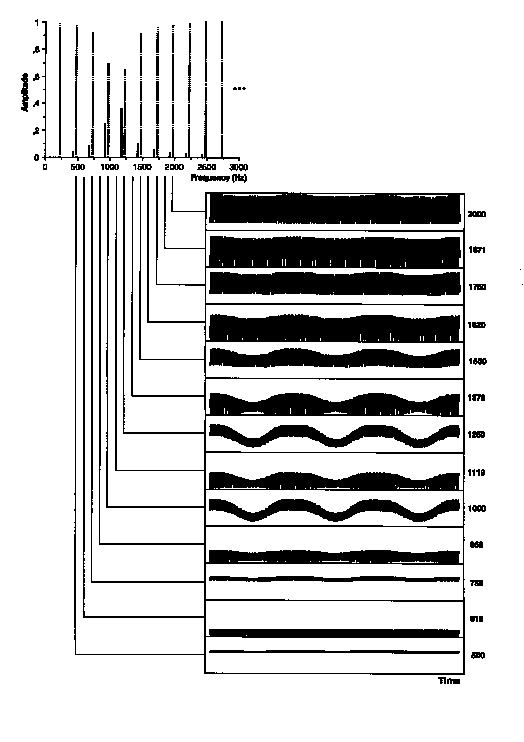
Figure 3. Example of envelopes of the output from gammatone filtering of a stimulus in Experiment 1: a frequency modulated test sound with a normal formant centered on 1150 Hz and an rms modulation width giving threshold matching (0.7%). The waveforms represent the envelope of the auditory filter output from 200 to 800 ms in the 1-s stimulus. The auditory filter CFs are listed to the right and their placement with respect to the stimulus components is indicated. Test components (solid) have been displaced with respect to background components (dotted) for visibility.
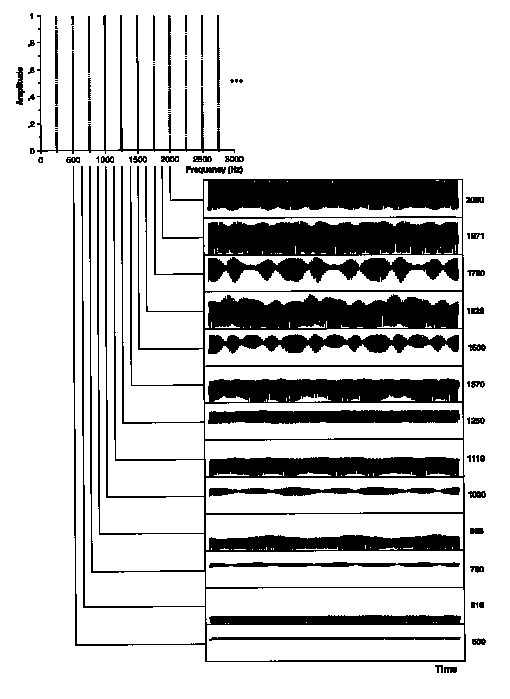
Figure 4. Example of envelopes of the output from gammatone filtering of a stimulus in Experiment 2: a frequency modulated test sound with a flat formant centered on 1250 Hz and an rms modulation width giving threshold matching (1.1%). (See Figure 5 caption.)
To quantify the amount of modulation present in a given auditory channel, the rms modulation index (mrms) was computed as the rms amplitude of the envelope divided by its mean taken over the steady-state portion of the sound between 200 and 800 ms. This index gives a mean-independent measure of variation about the envelope mean that is comparable across different, arbitrarily complex modulation functions. To represent a sort of fluctuation profile, mrms was plotted as a function of filter CF on an ERB-rate scale. These profiles are shown in Figure 5(a) for the stimuli represented in Figures 3 and 4. Note that mrms is greatest in auditory filters centered between stimulus components. As can be seen in those figures, the preponderance of the fundamental frequency (250 Hz) in the envelope increases with increasing filter CF. A corresponding increase in mrms is apparent in Figure 5(a), particularly for auditory filters centered on stimulus components.
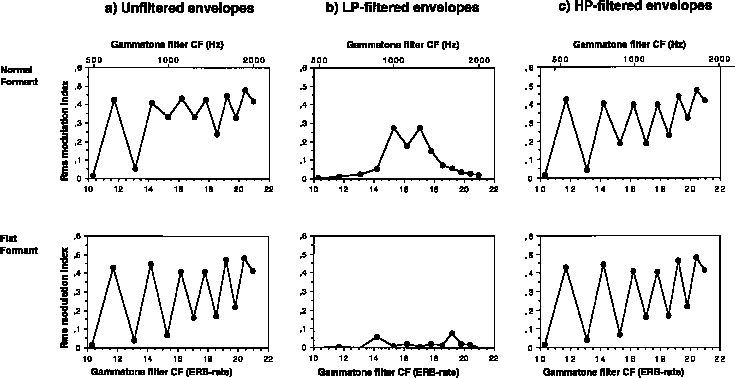
Figure 5. Fluctuation profiles (rms modulation index, mrms, as a function of auditory filter center frequency, ERB-rate) for the stimuli in Figures 3 and 4 derived from envelopes extracted from gammatone auditory filter output (a) and from the same envelopes that have been low-pass-filtered at 100 Hz (b) and high-pass filtered at 150 Hz (c).
Of greatest interest for our concerns are the envelope fluctuations in
the region of the formant skirts for normal formant stimuli or in the region
between adjacent test and background components in the flat format stimuli.
These fluctuations may result from several possible sources: 1) beating between
nearby test and background components for normal formant stimuli with
F0 or FM (Fig. 3), 2) induced AM as components move along the slopes of the formant skirts for normal formant stimuli with FM (Fig. 3), 3) induced AM in all FM stimuli as the modulated components move along the skirts of the auditory filters (Figs. 3 and 4), 4) beats of mistuned harmonic relations in regions between adjacent test and background components for flat formant
stimuli (Fig. 4), and 5) random envelope fluctuations due to the noise floor.
Of these five potential sources, the one most likely to be affected by our
choice of sine phase relations among stimulus components is the fourth. The
choice of random phases or other low peak-factor phase relations would decrease
the modulation depth due to this cue.
To examine more closely the nature of beats created in our stimuli, fluctuation profiles were created from the original envelope signals as well as from low-pass and high-pass versions of the envelopes. Low-rate components include sources 1-3 mentioned above, while source 4 is carried by higher frequencies in the vicinity of F0. Source 5 is a broader-band fluctuation source. A Kaiser filter with a 255-point impulse response and a stopband attenuation of 70 dB was applied to the extracted envelopes. A 100 Hz cut-off was used for the low-pass versions and a 150 Hz cut-off was used for the high-pass versions. The rms modulation index was then computed on these filtered envelopes. Since high-pass filtering removes the DC component, the envelope mean used for calculating mrms was taken from the unfiltered version. The result of the low-pass filtering process is shown in Figures 6 and 7 for the same stimuli in Figures 3 and 4.
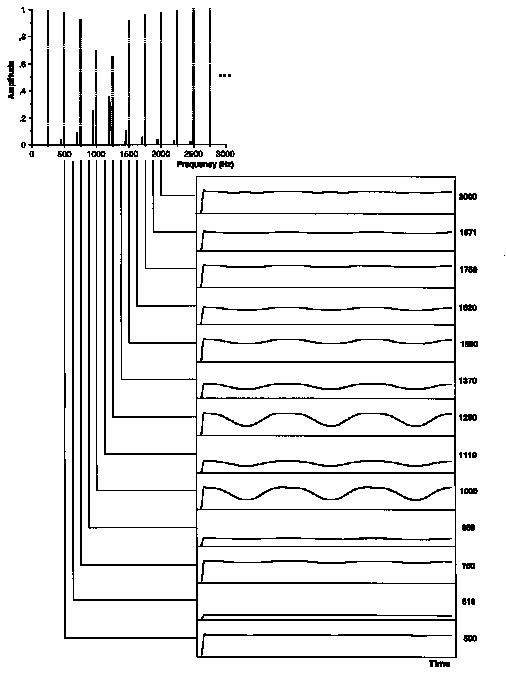
Figure 6. 100 Hz low-pass filtered envelopes for the stimulus represented in Figure 3.
Figure 7. 100 Hz low-pass filtered envelopes for the stimulus represented in Figure 4.
Of the five sources of low-rate beating mentioned above, this procedure preserves all but the fourth. Compare envelopes for the 1620 Hz channel between Figs. 4 and 7. In this channel, the beating in the unfiltered version is carried by higher periodicities interacting in the envelope that are attenuated by the filtering. To the contrary, beating due to nearby components in the 1250 Hz channel in Fig. 3 is preserved in Fig. 6. Note further that the low-rate fluctuations induced by FM in the skirts of auditory filters are very small at threshold FM widths (1119 and 1370 Hz channels in Fig. 7). The effects of filtering on the fluctuation profiles are shown in Figure 5(b,c). Note particularly that the modulation depth profile of lower-rate fluctuations signals the region of the normal formant but not the flat formant. The higher-rate fluctuations are prominent in both stimulus types. In general however, differences between the patterns for normal formants are most visible in profiles derived from low-pass filtered envelopes.
Results of the preceding experiments and model analyses suggest that, in some cases at least, beating in specific frequency regions may be used to perform the matching task. In these cases, low-rate sources of fluctuation seem to make the greatest distinction between test formants. Therefore, Experiment 6 sought to determine the amplitude modulation index necessary to just match the spectral region of auditory beats with a low-rate fluctuation pattern similar to those revealed by the above analyses, when this was the only cue available to perform the target matching task.
F0 stimuli depend on harmonic rank but have regions with the rates
included in this modulation waveform for all formant comparisons at threshold
except the highest CF formant in Experiment 4. The phase relations were
arbitrarily chosen from those found in the amplitude envelope of a frequency
modulated test formant with a 1700 Hz CF after passing through an auditory
filter centered on 1500 Hz. The AM was applied such that the mean level for
each component of the test formant over the duration of the tone corresponded
to the fixed level of the background components. This ensured that all test
stimuli had equal rms levels. For these stimuli, low-rate AM is localized only
in the region of the flat formant. The 250 Hz fluctuations are still present in
channels between lower harmonic components and in all higher-frequency
channels.A 16-turn adaptive tracking procedure was used with interleaved presentation of the three target CFs (375, 750, 1250 Hz). The dependent variable was the rms modulation index (mrms). This value varied from 0 to 0.3 in steps of 0.03. After training, five threshold estimates for each target formant were collected for two highly trained subjects (the authors).
More recently, Hall and Grose (1991) and Moore and Bacon (1993) have investigated subjects' abilities to identify which frequency component in a complex sound (two or six harmonic components) was sinusoidally amplitude modulated. Hall and Grose presented two-component signals with frequencies of 1 and 2 kHz and asked subjects to identify which component was amplitude modulated. Thresholds gave mrms values of 0.05 to 0.27. Moore and Bacon found that subjects' identification performance was well above chance with mrms values of 0.35 and 0.71, when a probe component was presented either before or after the complex signal containing a single modulated harmonic. The values from both of these studies agree with our thresholds. In addition, the Moore and Bacon tested identification of single, modulated components in 6-component complexes. Threshold mrms values varied from 0.14 to 0.22, indicating that sensitivity may be greater to the "odd man out" in multi-component backgrounds than in the presence of a single unmodulated component. The authors suggest that the modulation of a single component may promote its perceptual segregation, but that it is more difficult to then decide which component was modulated in the two-component complex than in the 6-component complex.
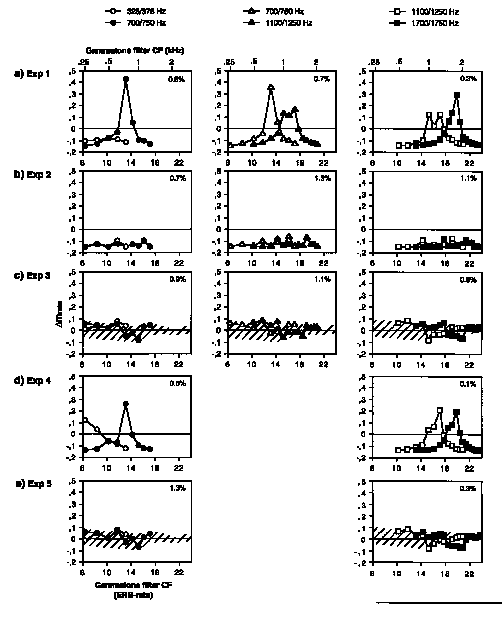
Figure 8. Differences in rms modulation index
(mrms)between threshold stimuli in Experiments 1-5 and those in Experiment 6 (with or without a pink noise floor as appropriate) as measured from model output. The difference is plotted as a function of the CF of the gammatone filter. Pairs of test formants are presented in columns and
experiments are presented in rows (a-e). The mrms values (see text) are
computed on low-pass filtered envelopes for gammatone filters centered on or
between harmonics in the frequency region around the test formant. The textured
region indicates the range of variation in mrms across the ten different
pink noise samples used in Experiments 3 and 5. The values in the upper left
hand corner of each panel are the threshold rms FM width or F0 used in the analysis.
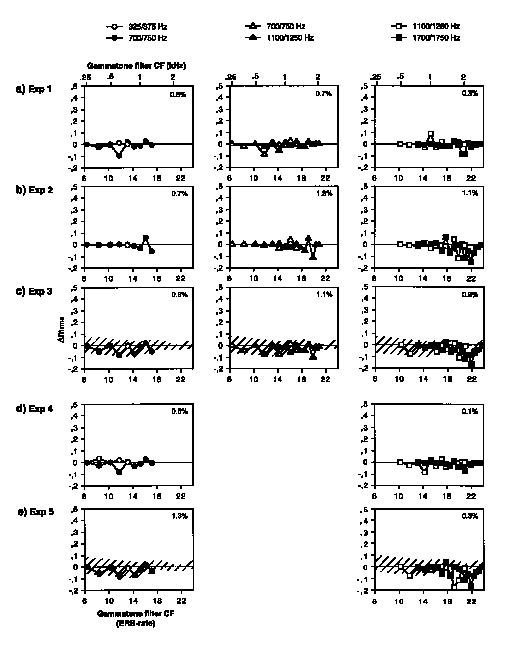
Figure 9. mrms for threshold stimuli in Experiments 1-5 computed on high-pass filtered envelopes (see Fig. 8 caption).
Figures 8 and 9 present the mrms values computed on low-pass and
high-pass filtered envelopes, respectively, at each CF for a given test formant
in each experiment after the corresponding values derived from threshold
stimuli in Experiment 6 have been subtracted (mrms). Thus,
values below zero are interpreted as being too small to allow formant matching
on the basis of this cue alone. If mrms is greater than zero,
this cue may be useful if the fluctuation profiles for the two formants are
sufficiently different. For stimuli from Experiments 1, 2, and 4 (quiet) the
value 0.15 was simply subtracted at all CFs since this threshold was nearly
constant across test formants. Profiles from the stimuli of Experiments 3 and 5
(pink noise floor) were derived by subtracting the corresponding profiles for
each threshold test stimulus in Experiment 6 presented against a noise floor.
This latter computation is not totally adequate since the profile would vary in
detail on a stimulus-to-stimulus basis as the noise floor waveform varied.
Recall that ten separate noise samples were used and were randomly selected for
each stimulus presentation. In order to give a sense of the possible effect of
the different noise samples, the range of variation in mrms
across the ten noise waveforms is indicated as the textured region in Figures
8(c) (Exp. 3) and 8(e) (Exp. 5). For our interpretations below, we consider
points lying within this noise range to be unreliable for performing the task
since they could be confounded with those obtained from the noise floor in the
absence of any harmonic stimulus. Further, this representation probably
overestimates sensitivity to modulation depth for higher-rate fluctuations
which Viemeister (1979) has shown to be less than that for low-rate
modulation.
It is of particular interest to compare the figures for FM or F0
across conditions expected to reduce beating [Figs. 8(a-c) and (d-e),
respectively]. For normal-formant FM stimuli at threshold (Exp. 1), one or both
formants create a significant amount of AM in the channels centered near the
test and background components that are nearly equal in amplitude. The channels
in which this AM occurs are different for each formant and discrimination of
these fluctuation patterns is a likely cue for target matching. A similar
explanation seems tenable for normal-formant F0 stimuli (Exp. 4). Note
that for threshold performance for both F0 and FM,
mrms is largely above zero, being greater than 0.2 for at
least one of two formants in all cases. This suggests that the effectiveness of
a pure modulation index-related cue is less in these stimuli than in simple AM
stimuli.
Now we can contrast these curves with those representing the envelope
fluctuations present in stimuli designed to reduce low-rate beating (Exps. 2,
3, and 5). Note that the mrms values are well below zero for
all formants in Experiment 2. This result would suggest that the beating that
is present is not useful for target matching. For stimuli with noise floors,
the mrms values vary around zero but rarely exceed the range
of variation due to the different noise samples. While this might suggest some
residual detectability, we feel we are justified in rejecting this hypothesis
for three reasons. Firstly, the actual pattern of variation differs from one
noise sample to the next and the variation between samples is approximately
equivalent to that found between the two formants being compared. As such, it
is difficult to imagine that the difference in fluctuation profile shown in
Figure 8 would allow reliable matching across noise samples. Secondly, for a
given noise sample, the mrms values do not increase monotonically with
the values of either dependent variable (rms modulation width or F0),
indicating that there is no reliable information in the depth of low-rate
fluctuation that would allow the subject to perform the task. Thirdly, the
differences in profile between pairs of test formants at threshold values of
the dependent variable are very small compared to those in threshold stimuli
from Experiments 1 and 4 [compare (a) with (c) and (d) with (e) in Fig. 8]. It
thus seems unlikely that detection of low-rate fluctuations was used to perform
the matching task for flat formant stimuli.
Nonetheless, closer examination of the unfiltered model outputs resulting from
flat-formant and flat-formant-plus-pink-noise stimuli reveals some apparent
periodic fluctuations in channels at and above the one centered on the highest
component of the test formant. In this region, there appears to be an
interaction between a 250 Hz periodicity resulting from overlapping background
components and either a slightly faster periodicity resulting from overlapping
test components in the F0 stimuli, or a separate periodicity that
modulates around this value resulting from overlapping test components in the
FM stimuli. Similar interactions between envelope periodicities are found on
the lower frequency side of higher-CF, flat-formant test sounds. In this way, a
lower-rate envelope fluctuation results from interaction of two higher-rate
envelope fluctuations, a phenomenon perhaps akin to beats of mistuned
consonances (Plomp,
1976).
Comparing fluctuation profiles for high-pass filtered envelopes (Fig. 9), it
becomes clear that sufficient mrms values are obtained for
threshold stimuli only in Experiments 1, 2, and 4. However, the differences
between profiles for the pair of test formants are very small in all cases. In
Experiments 3 and 5 (pink noise floors), the fluctuation profiles do not exceed
the variation due to the noise floor. So detection of modulation due to
higher-rate fluctuations would not appear to be a useful cue for formant
matching.
We cannot completely rule out, on the basis of these data, the possibility that the detection of differences in periodicity and shape of the higher-rate envelope fluctuations may have allowed detection of the frequency region of the test formant and thus its matching to the target formant in cases in which the modulation index itself did not distinguish between test formants. However, results from Experiment 6 suggest that detection or discrimination of such differences may not be sufficient in the present target matching paradigm. Further, work by Viemeister (1979) on temporal modulation transfer functions for broad-band noise suggests that sensitivity to modulation frequencies around 250 Hz is much less than that at low modulation frequencies. It seems likely that greater differences would be needed to perform this task.
2) Elimination of low-rate beats due to both interactions of nearby test and background components and tracing of formant spectral envelopes increased matching thresholds for higher-CF target formants that were modulated in frequency. Introduction of a noise floor gave no further increase in thresholds. Beating induced by frequency modulation of target components would thus seem to contribute significantly to target matching performance.
3) Target matching thresholds with fundamental frequency mistuning of normal formants are much lower than those found for vowel identification, most likely because a simple detection of the frequency region of beating between test formant and background would allow performance of this task. The particularly low threshold for higher CF formants (0.1% mistuning) suggests that subjects used fine-grained temporal information induced by the mistuning, although computer simulations suggest that the modulation depth itself is insufficient to explain this effect.
4) Elimination of low-rate beating and the addition of a noise floor
significantly increased F0 thresholds and all the more so for lower CF
targets. This increase at the lower CF target was much greater for F0
stimuli than for FM stimuli, suggesting that modulated targets create
additional, low-rate fluctuation cues not available with static targets in
stimuli with partially resolved harmonics. It is likely that this cue is
related to the AM induced in auditory filters by the FM.
5) Comparisons of experimental measures of threshold AM depth necessary to perform the matching task with measures of within-channel modulation index derived from computer simulations of auditory filtering suggested that low-rate beats contributed to performance with normal formants but were unlikely to have played a role for flat formant stimuli.
Therefore, low-rate beating in specific frequency regions is a likely cue for the perception of frequency modulated harmonic target sources that are embedded in an interfering harmonic background and whose spectral characteristics are known or predictable. One wonders whether this mechanism may not be responsible for the increased perceptual prominence found for frequency modulated vowels in multi-vowel mixtures that cannot be explained on the basis of segregation (Culling & Summerfield, 1995; McAdams, 1989). Further, low-rate beats are a likely cue for detecting embedded harmonic targets that are mistuned with respect to the background. However, in the absence of this cue, target sounds can still be perceived at sufficient mistuning from the background, suggesting additional contributions of either the pattern of higher-periodicity beating (mistuned consonances) and/or perceptual segregation on the basis of polyperiodicity itself.
1 If Experiment 1 thresholds are expressed as maximum percent mistuning attained by the FM waveform, the effect of cue type just misses attaining significance [F(1,4)=6.9, p=0.058]. No other differences occur between the two analyses.
2 For the 1250 Hz target in Experiment 5, the mean threshold for one subject was a factor of 8 (6.7 s.d.) higher than the mean of the other four subjects. All other subject means were within 1.5 s.d. of the group mean. As such we considered this data point an outlier and removed it from subsequent data analyses.
3 It is, of course, possible that the thresholds would not be identical in quiet and in noise. However, the spectra of the filtered envelopes for the AM stimuli in noise still reveal prominent components at the multiples of 5 Hz in channels centered on the modulated components.
Carlyon, R. P. (1991). "Discriminating between coherent and incoherent frequency modulation of complex tones," J. Acoust. Soc. Am. 89, 329-340.
Carlyon, R. P. (1992). "The psychophysics of concurrent sound segregation," Phil. Trans. R. Soc. Lond. B 336, 347-355.
Chalikia, M. H. and Bregman, A. S. (1989). "The perceptual segregation of simultaneous auditory signals: Pulse train segregation and vowel segregation," Percept. Psychophys. 46, 487-496.
Culling, J. F. and Darwin, C. J. (1993). "Perceptual separation of simultaneous vowels: within and across-formant grouping by F0," J. Acoust. Soc. Am. 93, 3454-3467.
Culling, J. F. and Summerfield, Q. (1995). "The role of frequency modulation in the perceptual segregation of concurrent vowels," J. Acoust. Soc. Am. 98, 837-846.
Cutting, J. E. (1976). "Auditory and linguistic processes in speech perception: inferences from six fusions in dichotic listening," Psychol. Rev. 83, 114-140.
de Cheveigné, A. (1993). "Separation of concurrent harmonic sounds: Fundamental frequency estimation and a time-domain cancellation model of auditory processing," J. Acoust. Soc. Am. 93, 3271-3290.
de Cheveigné, A., McAdams, S., Laroche, J., and Rosenberg, M. (1995). "Identification of concurrent harmonic and inharmonic vowels: A test of the theory of harmonic cancellation and enhancement," J. Acoust. Soc. Am. 97, 3736-3748.
Demany, L. and Semal, C. (1989). "Detection thresholds for sinusoidal frequency modulation," J. Acoust. Soc. Am. 85, 1295-1301.
Gardner, R. B. and Darwin, C. J. (1986). "Grouping of vowel harmonics by frequency modulation: Absence of effects on phonemic categorization," Percept. Psychophys. 40, 183-187.
Glasberg, G. R. and Moore, B. C. J. (1990). "Derivation of auditory filter shapes from notched-noise data," Hearing Res. 47, 103-138.
Goldstein, J. L. (1967). "Auditory nonlinearity," J. Acoust. Soc. Am. 41, 676-689.
Hall, J. W. and Grose, J. H. (1991). "Some effects of auditory grouping factors on modulation detection interference (MDI)," J. Acoust. Soc. Am. 90, 3028-3035.
Hartmann, W. M. and Klein, M. A. (1981). "The effect of uncertainty on the detection of frequency modulation at low modulation rates," Percept. Psychophys 30, 417-424.
Hartmann, W. M., McAdams, S., and Smith, B. K. (1990). "Hearing a mistuned harmonic in an otherwise periodic complex tone," J. Acoust. Soc. Am. 88, 1712-1724.
Helmholtz, H. L. F. (1885). On the Sensations of Tone as a Physiological Basis for the Theory of Music English trans. by A. J. Ellis from 4th German ed. (1877); republ. 1954 (Dover, New York).
Lea, A. (1992). "Auditory models of vowel perception," Unpublished doctoral dissertation, University of Nottingham, UK.
Levitt, H. (1971). "Transformed up-down methods in psychoacoustics," J. Acoust. Soc. Am 49, 467-477.
Marin, C. M. H. (1991). "Processus de séparation perceptive des sources sonores simultanées," Unpublished doctoral dissertation, Université de Paris III, France.
Marin, C. M. H. (1995). "Is fundamental frequency separation or harmonic coincidence a cue contributing to concurrent sound segregation?," (submitted).
Marin, C. M. H. and McAdams, S. (1990). "Segregation of concurrent sounds. II: Effects of spectral envelope tracing, frequency modulation coherence and frequency modulation width," J. Acoust. Soc. Am. 89, 341-351.
McAdams, S. (1984). "Spectral Fusion, Spectral Parsing, and the Formation of Auditory Images," Unpublished Ph.D. dissertation, Stanford University, Stanford, CA.
McAdams, S. (1989). "Segregation of concurrent sounds. I: Effects of frequency modulation coherence," J. Acoust. Soc. Am. 86, 2148-2159.
McAdams, S. and Marin, C. M. H. (1990). "Auditory processing of frequency modulation coherence", in Proceedings of the 6th Ann. Meeting Intl. Soc. Psychophys., Würzburg, pp. 175-180
Moore, B. C. J. and Bacon, S. (1993). "Detection and identification of a single modulated carrier in a complex sound," J. Acoust. Soc. Am. 94, 759-768.
Moore, B. C. J., Peters, R. W., and Glasberg, B. R. (1985). "Thresholds for the detection of inharmonicity in complex tones," J. Acoust. Soc. Am. 77, 1861-1867.
Patterson, R., Robinson, K., Holdsworth, J., McKeown, D., Zhang, C., and Allerhand, M. (1992). "Complex sounds and auditory images," in Auditory Physiology and Perception, edited by Y. Cazals, L. Demany, and K. Horner (Pergamon, Oxford), pp. 429-446.
Patterson, R. D. and Holdsworth, J. (1990). "An Introduction to Auditory Sensation Processing," (unpublished manual), MRC Applied Psychology Unit, Cambridge.
Plomp, R. (1976). Aspects of Tone Sensation: A Psychophysical Study trans. by from ed. republ. (Academic, London).
Riesz, R. R. (1928). "Differential sensitivity of the ear for pure tones," Phys. Rev. 31, 867-875.
Scheffers, M. T. M. (1983). "Sifting Vowels: Auditory Pitch Analysis and Sound Integration," Unpublished doctoral dissertation, University of Groningen, The Netherlands.
Smith, B. K. and Chervin, P. (1986). "Boris: An application of the Fujitsu MB8754 DSP chip", in Proceedings of the 1986 Intl. Comp. Mus. Conf., Den Haag, (Computer Music Association, San Francisco).
Summerfield, Q. and Assmann, P. F. (1991). "Perception of concurrent vowels: effects of harmonic misalignment and pitch-period asynchrony," J. Acoust. Soc. Am. 89, 1364-1377.
Summerfield, Q. and Culling, J. F. (1992). "Auditory segregation of competing voices: absence of effects of FM or AM coherence," Phil. Trans. R. Soc. Lond. B 336, 357-366.
Vercoe, B. (1986). Csound: A Manual for the Audio Processing System and Supporting Programs trans. by from ed. republ. (Media Lab, MIT, Cambridge, MA).
Viemeister, N. F. (1979). "Temporal modulation transfer functions based upon modulation thresholds," J. Acoust. Soc. Am. 66, 1364-1380.
____________________________
Server © IRCAM-CGP, 1996-2008 - file updated on .
____________________________
Serveur © IRCAM-CGP, 1996-2008 - document mis à jour le .